Hardy-Weinberg-Gleichgewicht
[Genfrequenz | Idealpopulation | HW-Gleichgewicht | Erweiterung]
Genfrequenz
Die simpelste Rechnung geht von 2 Allelen aus, deren
Häufigkeit im Folgenden untersucht werden soll. Das erste Allel sei A und das zweite a.
Gehen wir einmal davon aus, dass die Population, in der diese Allele vorkommen, aus diploiden
Individuen besteht. Dann trägt jedes Individuum zwei Allelenpaare. A und a können somit
in drei genotypischen Kombinationen auftreten: A+A, A+a, a+a. A+a ist "doppelt"
vorhanden wegen der zwei möglichen Kombinationen Aa und aA, die aber den gleichen
Phänotyp darstellen.
Um auf die jeweiligen Allelhäufigkeiten zu schliessen, muss man die Zahl der Individuen mit
AA, aa und 2xAa bestimmen. D sei die Anzahl der AA-homozygoten Individuen, H die Anzahl der
Aa-heterozygoten und R die Anzahl der aa-homozygoten Individuen.
Insgesamt hat die Population N Individuen, also:
D + H + R = N
Wenn z.B. bei Kaninchen die Haarfarbe bestimmt wird durch die Allele A für schwarze Haare
und a für weisse Haare, und die Allele kodominant sind, ist D die Anzahl der
schwarzhaarigen, R die Anzahl der weisshaarigen und H die Anzahl der grauhaarigen
Kaninchen in der Population der Grösse N. (Wenn A dominant und a rezessiv ist, ist R weiterhin
die Anzahl der weisshaarigen Kaninchen. Bei den schwarzhaarigen Kaninchen dagegen ist, nach den
Mendelschen Gesetzen zur dominant-rezessiven Vererbung, D 1/3 und H 2/3 der Anzahl der schwarzen
Kaninchen.)
In der diploiden Population kommt das Allel A genau 2D+H mal (wegen AA+Aa; D+0,5H) vor, a wiederum
2R+H mal (wegen Aa+aa; R+0,5H). Die Gesamtzahl aller Allele ist die doppelte
Populationsgrösse, also 2N. Deshalb erhält man die Häufigkeiten:
p[A] = Anz. A / Gesamtanz.Allele = (2D+H) / 2N = (D+.5H) / N
q[a] = Anz. a / Gesamtanz.Allele = (H+2R) / 2N = (.5H+R) / N
Wenn p schon berechnet ist, kann man auch mit q = 1 - p
die Häufigkeit des a-Allels berechnen.
Die Idealpopulation
Die Idealpopulation ist ein künstliches und stark
einschränkendes Populationsmodell das in den HW-Berechnungen verwendet wird. Die
Idealpopulation erfüllt folgende Bedingungen:
- die Population ist sehr (besser: unendlich) gross
- es herrscht Panmixie, d.h. die Individuen können sich
uneingeschränkt und mit gleicher Wahrscheinlichkeit paaren
- der Selektionsdruck ändert sich nicht
- es kommen keine Mutationen vor
- es tritt kein Gendrift, keine Populationsabspaltung und
keine Genmigration ein
Eine reelle Population erfüllt nicht alle dieser
Voraussetzungen. Deshalb liefern die folgenden Gesetze auf eine normale Population
angewandt keine exakten Ergebnisse.
Hardy-Weinberg-Gleichgewicht
G.H. Hardy, W. Weinberg und S. Tschetverikov entwickelten
zufällig zur gleichen Zeit, im Jahr 1908, Theorien zur Häufigkeit eines gewählten
Allels in den Filialgenerationen. Die nach den gefundenen Gesetzmässigkeiten
aufgestellten Formeln sind zusammengefasst unter dem Namen
"Hardy-Weinberg-Gleichgewicht". Tschetverikov wurde vergessen -
"Hardy-Weinberg-Tschetverikov-Gleichgewicht" als Benennung wäre wohl zu
lang gewesen.
Das HW-Gleichgewicht lässt sich folgendermassen berechnen: Wir nehmen eine
Idealpopulation, in der das Allel A mit der Frequenz p=P(A) und das Allel a mit der
Frequenz q=P(a) vorkommt. Aus D+H+R=N erhalten wir nach Teilen durch N den relativen
Anteil der drei Phänotypen in der Population.
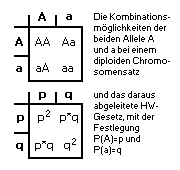 (D + H + R) / N = 1 (D + H + R) / N = 1
(Anz_AA+Anz_Aa+
Anz_aA+Anz_aa)/N = 1
also "AA + 2Aa + aa = 1".
mit "A"=P(A) und "a"=P(a)
=> P(A)2 + 2*P(A)*P(a) + P(a)2 = 1
oder mit P(a)=p und P(a)=q
=> p2 + 2pq + q2 = (p+q)2
= 1
Wenn die Voraussetzungen der Idealpopulation gelten, stellt die oben genannte Formel den
Gleichgewichtszustand da. Die Häufigkeiten p und q ändern sich nicht.
Wenn eine Population anfänglich nicht im Gleichgewichtszustand ist, stellt sich dieser
durch Einsetzen der Anfangswerte ein.
Zum Beispiel: eine sich nicht im Gleichgewicht befindende Population mit den
relativen Individuenhäufigkeiten
D1/N = 0,10 ; H1/N = 0,20 ; R1/N
= 0,70 und p = 0,20 ; q = 0,80
sieht in der F1-Generation folgendermassen aus
D2/N = p2 = 0,04 ; H2/N = 2pq =
0,32 ; R2/N = q2 = 0,64
Es ist D2/N + H2/N + R2/N = 1, also befindet sich die
Population in der F1-Generation im Gleichgewicht.
Mathematisch auffallend ist bei der Formel für den Gleichgewichtszustand auch,
dass die Häufigkeit der Heterozygoten nicht grösser als 0,5 sein kann. Die maximale
Anzahl an Heterozygoten wird erreicht bei p = q = 0,5
p2 + 2pq + q2 = (0,5)2 + 2*0,5*0,5 + (0,5)2
= 1
0,25 + 0,50 + 0,25 = 1
2pq = 2p(1-p) ; p<1 wird maximal, wenn 1-p=p also p=0,5
Um festzustellen, ob sich eine Population im Gleichgewichtszustand befindet, kann man
das folgende Gesetz anwenden:
=> 4*D*R = H2
z.B. 4 * 0,25 * 0,25 = 0,502
Weiterführung des HW-Gesetzes
In dem erweiterten Modell von Hardy und Weinberg wird die Häufigkeit eines Allelenpaars
(z.B. von aa) über mehrere Generationen hinweg untersucht. Bei dem Generationswechsel
wirkt auf aa ein konstanter Selektionsdruck, der mathematisch
dargestellt wird als Selektionsfaktor s. Der Selektionsfaktor liegt im Bereich von -1 bis
+1. Wenn s negativ ist, ist die Selektion für das untersuchte Allel günstig. Wenn s
positiv ist, hat das Allel einen Selektionsnachteil und die Häufigkeit des Allels in der
Population geht im Laufe der Generationen zurück.
Der Faktor s hängt von der relativen Fitness, d.h. von der relativen Anzahl der von aa
weitergegebenen Gene ab. Wenn die aa- und Aa-Genotypen 100% ihrer Allele, der AA-Genotyp
aber nur 20% weitergeben kann, entspricht das einer Fitness von wAA=0,2 und
einem Selektionsnachteil s=1-0,2=+0,8. Die Häufigkeit des A-Allels nimmt also ab,
während a zunimmt. (Graph unten).
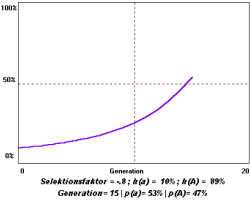 Die Formel für die
relative Häufigkeit des A-Allels ist rekursiv und etwas
komplizierter als die bisherigen Formeln: Die Formel für die
relative Häufigkeit des A-Allels ist rekursiv und etwas
komplizierter als die bisherigen Formeln:
q1 = (q0 - s * q02 ) / (1 - s * q02
)
...
qn = (qn-1 - s * qn-12
) / (1 - s * qn-12 )
Dabei ist qn-1 die relative Häufigkeit
des a-Allels in der untersuchten Generation. Die Formel gibt die Häufigkeit qn
für die nächste Generation an.
Wenn man die relative Häufigkeit von A nach zwei Generationen wissen will (q0
und s werden als bekannt vorausgesetzt), berechnet man q1, setzt das Ergebnis erneut
in die Formel ein und erhält das gesuchte q2. Die Häufigkeiten berechnet man
am besten mit einem Simulationsprogramm.
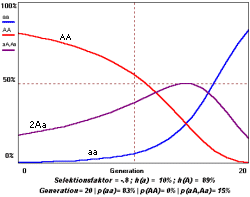 Wenn man q bestimmt hat,
kann man p leicht erhalten mit p=1-q. Mit diesen Werten kann man die relative Häufigkeit
der drei Phänotypen mit AA, Aa, aa bestimmen. Wenn man q bestimmt hat,
kann man p leicht erhalten mit p=1-q. Mit diesen Werten kann man die relative Häufigkeit
der drei Phänotypen mit AA, Aa, aa bestimmen.
Da p und q die Gleichung p2 + 2pq + q2 erfüllen, ist D / N =p2
; H / N =2pq ; R / N = q2. Ein Graph, in dem die Rekursionsformel für einige
Generationen durchgerechnet und mit p und q die Frequenz der Phänotypen bestimmt wurde,
könnte ähnlich aussehen wie oben. Die 50%-Grenze von den Aa-Genotypen ist hier gut
erkennbar.
Wie vielleicht schon aus der rekursiven Formel zu erkennen ist, hat dieses Modell die
weitere Eigenschaft, dass keines der Allele je die Häufigkeit von 100% bzw. 0% erreicht.
Diese Häufigkeiten sind Grenzwerte der Formel und werden innerhalb des Modells nie
erreicht. In der Natur dagegen kann, weil die Population endlich gross ist, ein Allel 100%
erreichen.
Zuletzt noch eine kleine Anmerkung: Das Grundmodell ist nicht auf zwei Allele beschränkt,
sondern kann auf beliebig viele Allele erweitert werden. Für drei Allele gilt (p+q+r)2=1
und für vier Allele entsprechend (p+q+r+s)2=1. Weitere
Informationen sind zu finden auf der Linkseite.
Die oben gezeigte Rekursionsformel lässt ausser Betracht, dass die aa- und Aa-Gentypen
auch eine unterschiedliche Fitness w haben können. Wenn die Fitness mitgerechnet wird,
erhält man als Grundgesetz wAAp2 + wAa2pq + waaq2
= 1.
(C) 1998 Florian Wagner
e-mail petwag@kolumbus.fi
|
